

| アカガイ 足筋 |
イガイ 閉殻筋 |
ホタテガイ 閉殻筋 |
マガキ 軟体部 |
ウバガイ 筋肉部 |
セタシジミ 軟体部 |
ヤマトシジミ 軟体部 |
ハマグリ 軟体部 |
アサリ 軟体部 |
タイラギ 閉殻筋 |
|
| タウリン | 389.3 | 984.0 | 1190.4 | 725.0 | 757.9 | 1.6 | 1.8 | 464.2 | 664.2 | 934.0 |
| アスパラギン酸 | 61.0 | 26.0 | 1.3 | 43.2 | 10.7 | 12.6 | 2.7 | 21.2 | 20.9 | 44.0 |
| トレオニン | 19.7 | 22.0 | 15.7 | 10.3 | 23.1 | 6.0 | 5.1 | 7.2 | 12.5 | 10.9 |
| セリン | 6.2 | 40.0 | 16.6 | 35.3 | 36.5 | 5.9 | 2.9 | 7.8 | 23.6 | 4.5 |
| グルタミン酸 | 129.5 | 69.0 | 108.8 | 142.0 | 156.0 | 21.9 | 23.5 | 76.9 | 102.8 | 85.0 |
| プロリー | 50.5 | ND | 44.5 | 55.0 | 26.1 | 6.7 | 21.9 | 8.0 | 16.4 | 20.8 |
| グリシン | 237.9 | 313.0 | 1985.0 | 242.0 | 458.9 | 8.8 | 5.1 | 86.4 | 328.5 | 136.7 |
| アラニン | 59.5 | 180.0 | 293.8 | 120.0 | 822.9 | 27.7 | 83.6 | 295.3 | 129.5 | 165.7 |
| シスチン | ND | ND | 1.2 | ND | trace | trace | ND | trace | 4.5 | ND |
| バリン | 5.3 | 4.0 | 5.3 | 5.2 | 12.3 | 5.4 | 5.5 | 6.7 | 14.4 | 2.8 |
| メチオニン | 15.7 | 7.0 | 2.4 | 2.3 | 5.8 | 5.4 | 0.6 | 4.2 | 10.5 | 2.5 |
| イソロイシン | 4.1 | 3.0 | 2.7 | 4.7 | 7.2 | 4.5 | 4.8 | 5.0 | 10.5 | 2.8 |
| ロイシン | 4.3 | 4.0 | 4.1 | 7.0 | 7.7 | 6.6 | 6.2 | 7.2 | 20.1 | 3.7 |
| チロリン | 9.0 | 17.0 | 2.6 | 5.6 | 19.8 | 6.5 | 3.0 | 7.1 | 16.3 | 1.9 |
| フェニルアラニン | 5.3 | 7.0 | 2.3 | 3.7 | 5.0 | 10.9 | 1.8 | 5.4 | 19.5 | 2.5 |
| β‐アラニン | 913.7 | 51.0 | 1.7 | 151.0 | - | - | 3.2 | - | - | 570.3 |
| トリプトファン | ND | 5.0 | 0.5 | ND | ND | 1.8 | trace | trace | ND | ND |
| ヒスチジン | 4.1 | 12.0 | 3.2 | 6.9 | 7.4 | 2.3 | 1.6 | 3.6 | 8.7 | 8.6 |
| リシン | 6.7 | 10.0 | 3.7 | 7.5 | 17.1 | 15.2 | 2.7 | 12.6 | 25.0 | 10.2 |
| アルギニン | 143.7 | 140.0 | 94.6 | 86.0 | 288.7 | 14.3 | 8.4 | 81.2 | 93.5 | 365.5 |
| 遊離アミノ酸総量 | 2065.5 | 1894.0 | 2780.8 | 1689.9 | 2663.1 | 164.1 | 185.5 | 1100.0 | 1521.3 | 2372.4 |
次に梅産の二枚貝をみると,総量で1100mg~3781.8mgと多く,上記のシジミ類との違いが明かである。また,マガキ,ハマグリおよびアサリに関しては,海産種の中では総量がそれぞれ1689,1100,1521mgと少なく,他の海産二枚貝と異なる。このことは分析試料の部位が上記3種以外の二枚貝では筋肉部であるのに対し内臓,外套部,鰓など軟体部全体を試料に供している点に原顔があると考えられ,含量を論議する際は部位の違いについても考慮する必要がある(4-1参照)。さらに,ホタテガイ閉殻筋の総量が極めて高いのに気づく。この点はアカガイ,イガイ,マガキなどは広塩性(広い範囲の釜分濃度の変化に対しても生存可能な生物―潮間帯に生息している軟体動物など)であるのに対し,ホタテガイは狭塩性(狭い範囲の塩分濃度の変動にのみ生存可能な生物―外洋性の軟体動物など)であることが起因しているとも考えられ,塩分耐性や生息環境,あるいは運動性と遊離アミノ酸レベルの関係など,今後多くの点について検討する必要がある(4参照)。
個々の遊離アミノ酸を通覧すると,軟体部を試料とした総量の少ない3種(マガキ,ハマグリ,アサリ)ではタウリン,グリシン,アラニン,グルタミン酸,アルギニンおよびβ‐アラニン(マガキ)が多く,5ないし6成分で総量の86%以上を占めている。筋肉部を試料とした総量の多い5種でもタウリン,グリシンが多く,次いでアラニン,グルタミン酸,アルギニン含量が高く,全種に共通した特徴を示している。特に,ホタテガイ(一成分で総量の52%,以下同様)のグリシン,アカガイ(44%)ではβ‐アラニン,イガイ(52%)とタイラギ(39%)のタウリン,ウバガイ(31%)でアラニンが多く,それぞれ最高値を示しているのが注目される。その他のアミノ故ではアスパラギン酸,トレオニン,セリンおよぴプロリンが比較的多く,それら以外は10mg以下のものが大部分である。
このように,個々の遊離アミノ酸含量は二枚貝の種類によりかなり異なるが,タウリン,グリシン,アラニン,β-アラニン,アルギニン,グルタミン酸などの特定の成分に分布が偏るという点は各種とも共通している。また,上記6種の成分中,アルギニンを除いてグリシン,アラニン,グルタミン酸は非必須アミノ酸であり,タウリン,β-アラニンは生体のタンパク質を構成しないアミノ酸であるなど,多量に存在するものはほとんどが解糖またはTCAサイクルの中間生成物にアミノ基を付加して合成されるもの,あるいはα-アミノ酸代謝系の産物である点など代謝面にも共通性があり,極めて興味深い。
3-2.D-アミノ酸
タンパク質を溝成しているL-アミノ酸以外に非天然型のD一アミノ酸が水産動物に存在することほすでに知られているが10),ある種の二枚貝にD-アラニンが多量に存在し,浸透圧調節に重要な役割を担っていることが示されている。最近の研究を一部紹介する。
Matsushimaら11)は,日本産5種の二枚貝のD‐アラニン含量を測定し(表3),ヤマトシジミ,ハマグリ,アサリの鰓 外套膜,足筋に29.4~363.5mgを認めている。また,淡水で飼育したヤマトシジミを340mOsmの海水に移し,経時的に取り出し足筋中のアラニン含量の変動を調べている。その結果,D-アラニンはL-アラニンとともに顕著に増加すろことを示し,D-アラニンが浸透圧調節物質であることを認めている。
表3 数種二枚貝組織のD-とL-アラニン含量(mg/100g)
| 二枚貝 | 組織 | D-アラニン | L-アラニン |
| ムラサキガイ | 鰓 | ― | 33.9±2.7 |
| 外套部 | ― | 71.3±8.0 | |
| マガキ | 鰓 | ― | 56.1±1.8 |
| 外套部 | ― | 73.1±13.4 | |
| ヤマトシジミ (340mOsm 海水飼育) |
鰓 | 180.9±9.8 | 203.1±12.5 |
| 外套部 | 220.1±8.0 | 220.1±9.8 | |
| ハマグリ | 鰓 | 160.4±15.1 | 177.3±16.0 |
| 外套部 | 139.0±11.6 | 136.3±14.3 | |
| 足筋 | 363.5±13.4 | 376.9±20.5 | |
| アサリ | 鰓 | 29.4±4.5 | 46.3±5.3 |
| 外套部 | 48.7±0.9 | 56.7±4.5 | |
| 足筋 | 74.8±7.1 | 83.7±6.2 |
測定値は各4固体の平均値±標準偏差を示す。―:検出限界以下
FelbeckとWiley12)は,カリフォルニア産の二枚貝17種について,D体のアラニン,アスパラギン酸およびバリン含量を調べ,D‐バリンは検出されなかったが,D-アスパラギン敢は9種類に,D-アラニンはムラサキイガイを除く全種に検出されたと報告している。
最近,Watanabeら13)も11種の二枚貝におけるD-アラニンの分布を調べている。その結果,異歯目に属する二枚貝に多量検出され,ミルクイで,820mg,ウバガイで400mg,バカガイで370mg,ハマグリで235mgなどと多く,D‐アラニンの全アラニンに対する割合はいずれも50%を超え,L体よりも含量的に多いと報じている。
なお,表4に主なアミノ酸のL型とD型による味の差を示した。このようにD-アラニンをはじめD一アミノ酸はL-アミノ駿よウも破い甘妹を呈するものが多く,呈味試験を行う際にはその存在に注意する必要がある。
表4 L型(天然型),D型(非天然型)によるアミノ酸の殊の相違14)
| アミノ酸 | L型 | D型 | アミノ酸 | L型 | D型 |
| アラニン | 甘 | 強甘 | メチオニン | 苦 | 甘 |
| セリン | 微甘 | 強甘 | ヒスチジン | 苦 | 甘 |
| トレオニン | 微甘 | 弱甘 | チロシン | 微苦 | 甘 |
| バリン | 苦 | 強甘 | トリプトファン | 苦 | 強甘 |
| ロイシン | 苦 | 強甘 | オルニチン | 苦 | 微甘 |
| イソロイシン | 苦 | 甘 | アルギニン | 微苦 | 弱甘 |
3-3.ヌクレオチドとその関連物質
水童動物筋肉こ含まれる主なヌクレオチドは,アデノシン三リン変(ATP)とその関連物質である。ATPは,アデノシンニリン酸(ADP),アデノシンーリン故(A丸IP)を経て,魚頚の蒙合ではA九℡が混アミノ化されイノシン酸(IMP)となり,IMPからリン酸が脱離してイノシン(HxR)になり,さらにリボースが除かれてヒポキサンチン(Hx)になる分解経路をとる。それに対して,二枚貝などの無脊椎動物は,AMPが先に脱リン酸を受けて,アデノシン(AdR)を生成し,次いで,AdRの脱アミノ化が起こりHxRに至る経路をとる。このように魚類と二枚貝などの無脊椎動物では分解経路が異なるため,魚類ではIMPが多量に蓄積されるが,二枚貝ではAMPが蓄積される。また,それら多量に蓄積される成分は重要な呈味成分である。
Suwetjaら15)は氷蔵中の数種海産無脊椎動物に含まれるATP関連化合物含量の変動を調べている。表5に結果の一部を示す。死直後ではATPが多く、時間経過に伴いATPが減少し,AMPの蓄積が見られる。しかし,アカガイやアサリではIMPの蓄積も認められている。このように二枚貝の筋肉にも魚類筋肉と同じATP分解経路が存在すると考えられ,さらに詳細な検討を加える必要がある。なお,二枚貝の死後IMPが蓄積するという事実は呈妹性との関連でも重要な研究テーマになり得ると思われる。
表5 数種二枚貝筋肉氷蔵中のATPとその関連物質の含量変化(μmol.g,湿重量)
| 経過日数 | ATP | ADP | AMP | IMP | AdR | HxR | Hx | |
| アカガイ | 0 | 1.16 | 1.00 | 2.05 | 0.39 | 0 | 0 | 0 |
| 1 | 0.14 | 0.95 | 3.20 | 0.19 | 0 | 0 | 0 | |
| 5 | 0.06 | 0.60 | 2.67 | 0.78 | 0 | 0.08 | 0.05 | |
| ホタテガイ | 0 | 3.48 | 1.18 | 1.56 | 0 | 0 | 0* | |
| 1 | 0.92 | 1.17 | 1.52 | 0 | 0 | 2.31* | ||
| マガキ | 0 | 0.92 | 0.34 | 1.46 | 0.23 | 0.38 | 0.05 | |
| 1 | 0.13 | 0.60 | 1.62 | 0.33 | 0.26 | 0.07 | 0 | |
| ハマグリ | 0 | 0.15 | 0.56 | 1.19 | 0 | 0 | 0.04 | 0 |
| 1 | 0.11 | 0.38 | 1.25 | 0.05 | 0 | 0.12 | 0 | |
| アサリ | 0 | 1.21 | 1.07 | 1.05 | 0.71 | 0 | 0.15 | 0 |
| 1 | 0.48 | 1.15 | 1.44 | 0.74 | 0 | 0.12 | 0 | |
| 5 | 0.21 | 0.39 | 0.49 | 1.65 | 0 | 0.42 | 0.60 | |
*HxR+Hx
3-4.4級アンモニウム塩基
ベタイン類,トリメチルアミン(TMA)とそのオキシド(TMAO)が4級
アンモニウム塩基に含まれる。
まず,これまで水産物に検出されたベタイン寮の構造を図1に挙げた。こ
れら以外に溝造の類似したベタインが確認されている。
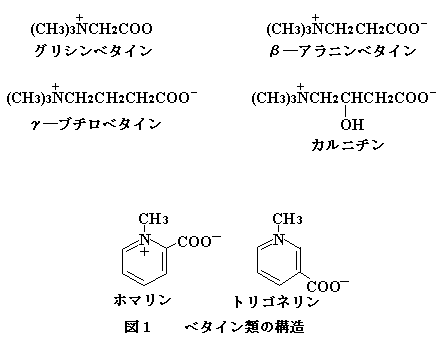
二枚貝中に含まれるベタイン類の含量を表6に示した。グリシンベタインおよびホマリンは海産二枚貝にかなり多量に含まれるのに対し,汽水産のヤマトシジミでは含量が極めて少ないことから浸透圧調節に寄与している考えられている。
表6 数種二枚貝筋肉のベタイン類含量*(mg/g)
| グリシンベタイン | ホマリン | |
| アカガイ | 900 | 101 |
| イガイ | 744 | 238 |
| ヒオウギガイ | 360 | 103 |
| マガキ | 1584 | 157 |
| ヤマトシジミ | 13 | 2 |
| ハマグリ | 493 | 69 |
| タイラギ | 1052 | 201 |
*品川ら 未発表データ.
なお,グリシンベタインほ甘味を有し,アワビでは重責な呈味成分であることが知られている16)ことから,二枚貝においても呈妹上の点でさらに追究する必要がある。
次にTMAとTMAOについて,数種二枚貝の含有量を表7に示した。サメ・エイなどの板鰓類に多いことはよく知られている9)が,二枚貝中にはイタヤガイやホタテガイに比較的多く存在する以外は少なく,淡水産のマシジミではほとんど検出されないことより,浸透圧調節に何らかの関与をしてい
ると考えられている。
表7 数種二枚貝のTMAO と TMA含量2,9) (mg/100g)
| TMAO | TMA | |
| イタヤガイ | 269 | 24 |
| ホタテガイ | 119 | 3 |
| ウバガイ | 11 | 1 |
| マシジミ | 1 | 0.5 |
なお,鮮度が低下すると腐敗臭を感ずるが,その原因物質は主にアンモニアとTMAで鮮度判定の指標としても使用されている。
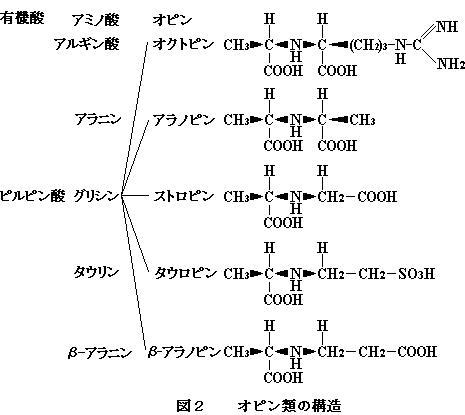
3-5.オピン済
オピン類はエキス成分の新顔として近年見いだされている。オピン類は分子内に共通してD-アラニンをもち,これと他のアミノ酸がイミノ基を共有する形で結合した構造をもつイミノ酸類の総称である。これまで二枚貝で確認されたものとしては,オクトビン,アラノピン,ストロンピン,タウロピンおよぴβ-アラノピンが知られている(図2)。これらはピルビン酸とアルギニン,アラニン,グリシン,タウリンおよびβ一アラニンなどがそれぞれ還元型ニコチンアミドアデニンジヌクレオチド(NADH)を補酵素として還元縮合することによって生成する。
一般に,二枚貝はかなりの嫌気耐性(低酸素の環境で生存し得る能力)を有する。ホタテガイを海水からとり空中に放置させると,中腸腺などの内臓組織にストロンピンの覇著な増加が認められるという。ムラサキイガイにおいてもアラニン,アラノピンまたはストロンピンなどの増加を認めている。さらに,アカガイでもβ-アラノピンが4℃,24時間の空中放置で急激に増加することを確認している17)。
このような二枚貝中のオピン類の研究により,オピン脱水素酵素類が脊椎動物の乳酸脱水素酵素と同義的な役乱すなわち嫌気的条件下におけるエネルギー獲得機構と深く関連していることが明らかになった。例示すれぼ,嫌気的解糖により生成したピルピン酸をオピン祭に変換すると同時に,解糖反応系で生じた補酵素NADHの再酸化を担い,NAD+を持続的に供給し,酸化還元平衝の維持をつかさどることになる。
3-6.糖および有機酸
糖,とりわけ二枚貝において重責な糖はエネルギー源としてのグリコーゲンであるぅ グリコーゲンの生理的意義としては解糖系やTCA回路を経るエネルギー産生メガニズムの起点としての役割および脂肪酸やアミノ酸合成素材としても重要な役割を有する。
二枚貝のグリコーゲン量を調べた例としては,高木ら18)の報告がある。彼らはムラサキイガイ,マガキ,マシジミおよびアサリについて水分量とグリコーゲン量の季節変動を調べている。その結果の一部を表8に示す。アサリほ季節変動は比較的少ないが,春から夏にかけて多くなる傾向を示している。マガキでは不味といわれる夏季に水分量の上昇とグリコーゲン量の急激な減少を認めている。これとは反対に,冬季では水分量の減少とグリコーゲン量の増加を認め,旬の時期にグリコーゲン量が増加するとしている。また,ムラサキイガイは夏,マシジミは冬が旬といわれているが,いずれもそれらの季節に水分が減少し,グリコーゲンがわずかながら増加している。
表8 二枚貝軟体部中の水分とグリコーゲン量の季節変動(%)
| 採取時期 | 水分量 | グリコーゲン量 | |
| ムラサキイガイ | 5月 | 81.8 | 1.25 |
| 9月 | 80.4 | 1.82 | |
| マガキ | 7月 | 85.0 | 1.30 |
| 12月 | 81.7 | 4.30 | |
| マシジミ | 6月 | 84.1 | 0.93 |
| 1月 | 83.1 | 1.00 | |
| アサリ | 6月 | 80.9 | 2.50 |
| 9月 | 82.2 | 1.90 | |
| 11月 | 78.8 | 1.00 | |
| 1月 | 83.0 | 1.50 | |
| 3月 | 79.1 | 2.40 |
有機酸,なかでも二枚貝では,コハク酸が嫌気的解糖系の最終産物およびうま味成分として重要である。二枚貝の有機酸を測定した報告例として鴻巣らの詳細な研究19)がある。彼らはアサリのコハク酸濃度が文献によって大きな差があることに疑問を呈し,試料の来歴を詳細に記した報告をしている。すなわち採取後から分析に供するまでの経過時間,採取した季節などが分析結果に大きく影響することを確かめている。その結果の一部を表9に示す。これによると4種の有機酸を定量し,最も多く検出される成分はコハク酸で,次いでリンゴ酸,フマル酸,乳酸の順で含量が低下している。コハク酸は12月のアサリで18mg,6月では40mgであるが,これをそれぞれ7℃および25℃に空中放置するとコハク酸のみが著しく増加する。特に夏季のアサリのコハク酸が冬季のアサリより含量がかなり高いことも報告している。
表9 アサリ採取後の時間経過に伴う有機酸含量 (mg/100g)
| 時期 | 経過時間 | フマル酸 | コハク酸 | リンゴ酸 | 乳酸 |
| 12月 (7℃貯蔵) |
0 | 2 | 19 | 17 | 2 |
| 10 | 1 | 30 | 20 | 3 | |
| 20 | 6 | 36 | 11 | 2 | |
| 32 | 4 | 34 | 5 | 2 | |
| 44 | 6 | 48 | 10 | 2 | |
| 88 | 2 | 60 | 16 | 2 | |
| 6月 (25℃貯蔵) |
0 | 10 | 40 | 12 | 5 |
| 8 | 8 | 84 | 9 | 1 | |
| 20 | 6 | 120 | 6 | 2 | |
| 32 | 6 | 170 | 4 | 3 | |
| 44 | 10 | 204 | 6 | 3 | |
| 56 | 5 | 238 | 14 | 4 |
また,山中ら20)はハマグリについて同様の実験を行い,2℃,2日間の空中放置で110mgと初濃度の約2倍の増加を示したとしている。他の二枚貝ではマシジミ 19)128mg(渓取後20~24時間経過,11月の試料),ホタテガイ19)14mg(採取後氷冷下30時間経過,12月の試料)などの判定結果がある。
3-7.無機質
二枚貝をはじめ,生物の体内には多くの元素が存在する。そのうち多量に存在する炭素,水素,酸素および窒素以外の元素は無機質と呼ばれる。無機質に分類される元素のうち仕較的多量に存在するのほ,ナトリウム,カリウム,カルシウム,マグネシウム,塩素,リン,イオウの7元素で,以上11元素で生体を構成している元素のほぼ100%を占める。これら以外の元素が微量元素と呼ぼれる。
二枚貝の無機質含量は,動物種や体内組織により異なり,特に貝殻では成
分のはとんどを占める。一方,筋肉などの組織では2%前後と低いが,タン
バタ質や脂質の構成成分として代謝のあらゆる面で重要な働きをしている。
また,体液中の無機質は主にイオンとして存在し,浸透圧調節や酸塩基平衡,
さらにタンパク質などの機能発現に関与しており,内部環境の恒常性を保つ
上で必須の成分である。
表10に分析データの多いナトリウム,カリウム,カルシウムおよびリンに
ついての二枚貝可食部中の含量を示す。
表10 二枚貝可食部の無機質含量 * (mg/100g)
| ナトリウム | カリウム | カルシウム | リン | |
| アカガイ | 300 | 290 | 40 | 140 |
| イガイ | 540 | 230 | 43 | 160 |
| マガキ | 280 | 230 | 55 | 130 |
| シジミ | 320 | 120 | 320 | 95 |
| ハマグリ | 500 | 250 | 140 | 110 |
| アサリ | 400 | 230 | 80 | 180 |
| タイラギ | 260 | 260 | 16 | 165 |
*四訂日本食品標準成分表より引用.
ナトリウム含量は300~540mgと魚顛70mgのなどに比べて多い傾向にある。主に細胞外液に存在し,浸透圧調節に関与している。カリウム含量は汽水産のヤマトシジミと思われるシジミに120mgと少なく,他の海産二枚貝では230~290mgと多い。ナトリウムとは反対に細胞内の主要無機質で,細胞内の浸透圧調節や筋肉収縮,神経興奮伝達など生理的意義を有する物質である。カルシウム含量はシジミに820mgと特に多く,他の二枚貝ではおおむね100mg以下と少ない傾向にある。カルシウムも血液凝固,筋肉収縮,神経の興奮調節などに関与し重要な無機質である。リンは種による変動は他の無機質に比べ少ない。主にリン酸イオンとして存在し,酸塩基平衡に重要な役割を担うとともに,核酸,リン脂質などの構成成分として有機リン酸エステルで存在し,さまざまな生体機能に関与している。
また,食品化学的こ見て,無機質は食品の特徴のある味を作り出すために不可欠な成分であることを付記する(5参照)。